MMP2
72 kDa type IV collagenase also known as matrix metalloproteinase-2 (MMP-2) and gelatinase A is an enzyme that in humans is encoded by the MMP2 gene.[5] The MMP2 gene is located on chromosome 16 at position 12.2.[6]



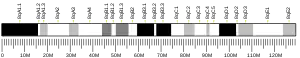
Function
Proteins of the matrix metalloproteinase (MMP) family are involved in the breakdown of extracellular matrix (ECM) in normal physiological processes, such as embryonic development, reproduction, and tissue remodeling, as well as in disease processes, such as arthritis and metastasis. Most MMP's are secreted as inactive proproteins which are activated when cleaved by extracellular proteinases. This gene encodes an enzyme which degrades type IV collagen, the major structural component of basement membranes. The enzyme plays a role in endometrial menstrual breakdown, regulation of vascularization and the inflammatory response.[7] The role of MMP2 in lymphangiogenesis was considered in a modelling and theoretical study; MMP2 degrades collagen I to switch on different patterning mechanisms for VEGFC.[8][9]
Activation
Activation of MMP-2 requires proteolytic processing. A complex of membrane type 1 MMP (MT1-MMP/MMP14) and tissue inhibitor of metalloproteinase 2 recruits pro-MMP 2 from the extracellular milieu to the cell surface. Activation then requires an active molecule of MT1-MMP and auto catalytic cleavage. Clustering of integrin chains promotes activation of MMP-2. Another factor that will support the activation of MMP-2 is cell-cell clustering. A wild-type activated leukocyte cell adhesion molecule (ALCAM) is also required to activate MMP-2.
Clinical significance
Mutations in the MMP2 gene are associated with Torg-Winchester syndrome, multicentric osteolysis, arthritis syndrome,[10] and possibly keloids.
Role of MMP-2 in chronic disease
Activity of MMP-2 relative to the other gelatinase (MMP-9) has been associated with severity of chronic airway diseases including Idiopathic interstitial pneumonia and Bronchiectasis. In idiopathic interstitial pneumonia, MMP-2 activity was elevated in patients with the less severe disease phenotype which is more responsive and reversible with corticosteroid therapy.[11] In non-cystic fibrosis bronchiectasis, MMP-2 concentration was elevated in patients with Haemophilus influenzae airway infection compared to Pseudomonas aeruginosa airway infection.[12] Bronchiectasis patients with P. aeruginosa infection have a more rapid decline in lung function.[13]
Altered expression and activity levels of MMPs have been strongly implicated in the progression and metastasis of many forms of cancer. Increased MMP-2 activity has also been linked with a poor prognosis in multiple forms of cancer including colorectal, melanoma, breast, lung, ovarian, and prostate.[14] Furthermore, changes in MMP-2 activity can come from alterations in levels of transcription, MMP secretion, MMP activation, or MMP inhibition. MMP production in many cancers may be upregulated in surrounding stromal tissue rather than simply in the tumor lesion. For instance, Mook, et al. showed that MMP-2 mRNA levels are strikingly similar between metastatic and non-metastatic lesions in colorectal cancer, but metastatic cases are correlated with higher levels of MMP-2 mRNA in surrounding healthy tissue.[15] For this reason, it is difficult to fully understand the complex role of MMPs in cancer progression.
Role in cancer cell invasion
One of the major implications of MMPs in cancer progression is their role in ECM degradation, which allows cancer cells to migrate out of the primary tumor to form metastases. More specifically, MMP-2 (along with MMP-9) is capable of degrading type IV collagen, the most abundant component of the basement membrane. The basement membrane is important for maintaining tissue organization, providing structural support for cells, and influencing cell signaling and polarity. Degradation of the basement membrane is an essential step for the metastatic progression of most cancers.[15]
Cancer cell invasion, ECM degradation, and metastasis are highly linked with the presence of invadopodia, protrusive and adhesive structures on cancer cells. Invadopodia have been shown to concentrate MMPs (including MT1-MMP, MMP-2, and MMP-9) for localized release and activation.[16] Furthermore, degradation products of MMP activity may further promote invadopodia formation and MMP activity.[17] Finally, MMP-2 and several other MMPs have been shown to proteolytically activate TGF-β, which has been shown to promote epithelial mesenchymal transition (EMT), a key process involved in cancer metastasis.[18]
Role in cell signaling
MMP degradation of the ECM affects cellular behavior through changes in integrin-cell binding, by releasing growth factors harbored by the ECM, by generating ECM degradation products, and by revealing cryptic binding sites in ECM molecules.[19] For instance, MMP-2 degradation of collagen type I can reveal a previously inaccessible cryptic binding site that binds with the αvβ3 integrin expressed by human melanoma cells. Signaling through this integrin is necessary for melanoma cell viability and growth in a collagen matrix and can potentially rescue the cells from apoptosis.[20] As another example, cleavage of laminin-5, a component of the basement membrane, by MMP-2 has been shown to reveal a cryptic site inducing migration of breast epithelial cells.[21]
More generally, by degrading the ECM, MMPs release growth factors that were previously bound to the ECM, allowing them to bind with cell receptors and influence cell signaling. Furthermore, many MMPs also activate other proMMPs along with growth factors.[19] MMP-2 has also been shown to cleave other non-ECM substrates including growth factors such as TGF-β, FGF receptor-1, proTNF, IL-1β and various chemokines.[22] For instance, MMP-2 has been implicated, along with MMP-9 in cleaving latent TGF-β, which has complex interactions with cancer cells. TGF-β generally plays a role in maintaining tissue homeostasis and preventing tumor progression. However, genetically unstable cancer cells can often evade regulation by TGF-β by altering TGF-β receptors in downstream signaling processes. Furthermore, expression of TGF-β is also correlated with immune tolerance and may help shield cancer cells from immune regulation.[23]
Role in neovascularization and lymphangiogenesis
MMP-2 also plays an important role in the formation of new blood vessels within tumors, a process known as angiogenesis. This process is essential for tumor progression, because as tumors grow they need increasing supplies of oxygen and nutrients. Localized MMP-2 activity plays an important role in endothelial cell migration, a key feature of angiogenesis. Additionally, MMP-9 and other MMPs have been suggested to also play a complex, indirect role in angiogenesis by promoting VEGF mobilization and generating antiangiogenic factors.[15]
For instance, when studying carcinogenesis of pancreatic islets in transgenic mice, Bergers et al. showed that MMP-2 and MMP-9 were upregulated in angiogenic lesions and that the upregulation of these MMPs triggered the release of bioactive VEGF, a potent stimulator of angiogenesis. Additionally, the group determined that MMP-2 knockout mice showed decreased rates of tumor growth relative to tumor growth rates in wild type mice.[24] Furthermore, increased expression and activity of MMP-2 has been tied to increased vascularization of lung carcinoma metastases in the central nervous system, which likely increases survival rate of these metastases.[25]
Finally, MMP-2 has been also shown to drive lymphangiogenesis, which is often excessive in tumor environments and can provide a route of metastasis for cancer cells. Detry, et al. showed that knocking down mmp2 in zebrafish prevented the formation of lymphatic vessels without altering angiogenesis, while MMP-2 inhibition slowed the migration of lymphatic endothelial cells and altered the morphology of new vessels.[15] These results suggest that MMP-2 may alter tumor viability and invasion by regulating lymphangiogenesis in addition to angiogenesis.
Inhibition of MMP-2 as cancer therapy
Clinical trials for cancer therapies using MMP inhibitors have yielded generally unsuccessful results. These poor results are likely due to the fact that MMPs play complex roles in tissue formation and cancer progression, and indeed many MMPs have both pro and anti-tumorogenic properties. Furthermore, most clinical studies involve advanced stages of cancer, where MMP inhibitors are not particularly effective. Finally, there are no reliable biomarkers available for assessing the efficacy of MMP inhibitors and MMPs are not directly cytotoxic (so they do not cause tumor shrinkage), so it is difficult for researchers to determine whether the inhibitors have successfully reached their targets.[14]
However, initial clinical trials using broad spectrum MMP inhibitors did show some positive results. Phase I clinical trials showed that MMP inhibitors are generally safe with minimal adverse side effects. Additionally, trials with marimastat did show a slight increase in survival of patients with gastric or pancreatic cancer.[14]
Various research groups have already suggested many strategies for improving the effectiveness of MMP inhibitors in cancer treatment. First, highly specific MMP inhibitors could be used to target the functions of specific MMPs, which should allow doctors to increase the treatment dosage while minimizing adverse side effects. MMP inhibitors could also be administered along with cytotoxic agents or other proteinase inhibitors. Finally, MMP inhibitors could be used during earlier stages of cancer to prevent invasion and metastasis.[14]
Additionally, tumor overexpression of MMPs can be used to potentially target the release of chemotherapeutic agents specifically to tumor sites. For instance, cytotoxic agents or siRNA could be encapsulated in liposomes or viral vectors that only become activated upon proteolytic cleavage by a target MMP. Finally, the tumor-targeting properties of MMP inhibitors offer a potential strategy for identifying small tumors. Researchers could couple MMP inhibitors to imaging agents to help detect tumors before they spread. Though initial trials yielded disappointing results, MMP inhibitors offer significant potential for improving cancer treatment by slowing the process of cancer cell invasion and metastasis.[14]
Interactions
MMP2 has been shown to interact with:
References
- GRCh38: Ensembl release 89: ENSG00000087245 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000031740 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Devarajan P, Johnston JJ, Ginsberg SS, Van Wart HE, Berliner N (December 1992). "Structure and expression of neutrophil gelatinase cDNA. Identity with type IV collagenase from HT1080 cells". J. Biol. Chem. 267 (35): 25228–32. PMID 1460022.
- "MMP2 gene". Genetics Home Reference. Retrieved 19 May 2015.
- "Entrez Gene: MMP2 matrix metallopeptidase 2 (gelatinase A, 72kDa gelatinase, 72kDa type IV collagenase)".
- Wertheim, Kenneth Y.; Roose, Tiina (April 2017). "A Mathematical Model of Lymphangiogenesis in a Zebrafish Embryo". Bulletin of Mathematical Biology. 79 (4): 693–737. doi:10.1007/s11538-017-0248-7. ISSN 1522-9602. PMC 5501200. PMID 28233173.
- Roose, Tiina; Wertheim, Kenneth Y. (3 January 2019). "Can VEGFC Form Turing Patterns in the Zebrafish Embryo?". Bulletin of Mathematical Biology. 81 (4): 1201–1237. doi:10.1007/s11538-018-00560-2. ISSN 1522-9602. PMC 6397306. PMID 30607882.
- Martignetti JA, Aqeel AA, Sewairi WA, Boumah CE, Kambouris M, Mayouf SA, Sheth KV, Eid WA, Dowling O, Harris J, Glucksman MJ, Bahabri S, Meyer BF, Desnick RJ (July 2001). "Mutation of the matrix metalloproteinase 2 gene (MMP2) causes a multicentric osteolysis and arthritis syndrome". Nat. Genet. 28 (3): 261–5. doi:10.1038/90100. PMID 11431697. S2CID 24810941.
- Suga M, Iyonaga K, Okamoto T, Gushima Y, Miyakawa H, Akaike T, Ando M (November 2000). "Characteristic elevation of matrix metalloproteinase activity in idiopathic interstitial pneumonias". American Journal of Respiratory and Critical Care Medicine. 162 (5): 1949–56. doi:10.1164/ajrccm.162.5.9906096. PMID 11069839.
- Taylor SL, Rogers GB, Chen AC, Burr LD, McGuckin MA, Serisier DJ (May 2015). "Matrix metalloproteinases vary with airway microbiota composition and lung function in non-cystic fibrosis bronchiectasis". Annals of the American Thoracic Society. 12 (5): 701–7. doi:10.1513/AnnalsATS.201411-513OC. PMID 25679336.
- Rogers GB, Zain NM, Bruce KD, Burr LD, Chen AC, Rivett DW, McGuckin MA, Serisier DJ (May 2014). "A novel microbiota stratification system predicts future exacerbations in bronchiectasis". Annals of the American Thoracic Society. 11 (4): 496–503. doi:10.1513/AnnalsATS.201310-335OC. PMID 24592925.
- Björklund M, Koivunen E (May 2005). "Gelatinase-mediated migration and invasion of cancer cells". Biochimica et Biophysica Acta (BBA) - Reviews on Cancer. 1755 (1): 37–69. doi:10.1016/j.bbcan.2005.03.001. hdl:10138/22049. PMID 15907591.
- Mook OR, Frederiks WM, Van Noorden CJ (December 2004). "The role of gelatinases in colorectal cancer progression and metastasis". Biochimica et Biophysica Acta (BBA) - Reviews on Cancer. 1705 (2): 69–89. doi:10.1016/j.bbcan.2004.09.006. PMID 15588763.
- Jacob A, Prekeris R (2015). "The regulation of MMP targeting to invadopodia during cancer metastasis". Frontiers in Cell and Developmental Biology. 3: 4. doi:10.3389/fcell.2015.00004. PMC 4313772. PMID 25699257.
- Clark ES, Whigham AS, Yarbrough WG, Weaver AM (May 2007). "Cortactin is an essential regulator of matrix metalloproteinase secretion and extracellular matrix degradation in invadopodia". Cancer Research. 67 (9): 4227–35. doi:10.1158/0008-5472.CAN-06-3928. PMID 17483334.
- Gialeli C, Theocharis AD, Karamanos NK (January 2011). "Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting". The FEBS Journal. 278 (1): 16–27. doi:10.1111/j.1742-4658.2010.07919.x. PMID 21087457. S2CID 2260074.
- McCawley LJ, Matrisian LM (April 2000). "Matrix metalloproteinases: multifunctional contributors to tumor progression". Molecular Medicine Today. 6 (4): 149–56. doi:10.1016/s1357-4310(00)01686-5. PMID 10740253.
- Montgomery AM, Reisfeld RA, Cheresh DA (September 1994). "Integrin alpha v beta 3 rescues melanoma cells from apoptosis in three-dimensional dermal collagen". Proceedings of the National Academy of Sciences of the United States of America. 91 (19): 8856–60. doi:10.1073/pnas.91.19.8856. PMC 44705. PMID 7522323.
- Giannelli G, Falk-Marzillier J, Schiraldi O, Stetler-Stevenson WG, Quaranta V (July 1997). "Induction of cell migration by matrix metalloprotease-2 cleavage of laminin-5". Science. 277 (5323): 225–28. doi:10.1126/science.277.5323.225. PMID 9211848.
- Detry B, Erpicum C, Paupert J, Blacher S, Maillard C, Bruyère F, Pendeville H, Remacle T, Lambert V, Balsat C, Ormenese S, Lamaye F, Janssens E, Moons L, Cataldo D, Kridelka F, Carmeliet P, Thiry M, Foidart JM, Struman I, Noël A (May 2012). "Matrix metalloproteinase-2 governs lymphatic vessel formation as an interstitial collagenase" (PDF). Blood. 119 (21): 5048–56. doi:10.1182/blood-2011-12-400267. PMID 22490679.
- Massagué J (July 2008). "TGFbeta in Cancer". Cell. 134 (2): 215–30. doi:10.1016/j.cell.2008.07.001. PMC 3512574. PMID 18662538.
- Bergers G, Brekken R, McMahon G, Vu TH, Itoh T, Tamaki K, Tanzawa K, Thorpe P, Itohara S, Werb Z, Hanahan D (October 2000). "Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis". Nature Cell Biology. 2 (10): 737–44. doi:10.1038/35036374. PMC 2852586. PMID 11025665.
- Rojiani MV, Alidina J, Esposito N, Rojiani AM (2010). "Expression of MMP-2 correlates with increased angiogenesis in CNS metastasis of lung carcinoma". International Journal of Clinical and Experimental Pathology. 3 (8): 775–81. PMC 2993228. PMID 21151391.
- McQuibban GA, Gong JH, Tam EM, McCulloch CA, Clark-Lewis I, Overall CM (August 2000). "Inflammation dampened by gelatinase A cleavage of monocyte chemoattractant protein-3". Science. 289 (5482): 1202–6. doi:10.1126/science.289.5482.1202. PMID 10947989.
- Bein K, Simons M (October 2000). "Thrombospondin type 1 repeats interact with matrix metalloproteinase 2. Regulation of metalloproteinase activity". J. Biol. Chem. 275 (41): 32167–73. doi:10.1074/jbc.M003834200. PMID 10900205.
- Morgunova E, Tuuttila A, Bergmann U, Tryggvason K (May 2002). "Structural insight into the complex formation of latent matrix metalloproteinase 2 with tissue inhibitor of metalloproteinase 2". Proc. Natl. Acad. Sci. U.S.A. 99 (11): 7414–9. doi:10.1073/pnas.102185399. PMC 124245. PMID 12032297.
- Overall CM, Tam E, McQuibban GA, Morrison C, Wallon UM, Bigg HF, King AE, Roberts CR (December 2000). "Domain interactions in the gelatinase A.TIMP-2.MT1-MMP activation complex. The ectodomain of the 44-kDa form of membrane type-1 matrix metalloproteinase does not modulate gelatinase A activation". J. Biol. Chem. 275 (50): 39497–506. doi:10.1074/jbc.M005932200. PMID 10991943.
- Bigg HF, Shi YE, Liu YE, Steffensen B, Overall CM (June 1997). "Specific, high affinity binding of tissue inhibitor of metalloproteinases-4 (TIMP-4) to the COOH-terminal hemopexin-like domain of human gelatinase A. TIMP-4 binds progelatinase A and the COOH-terminal domain in a similar manner to TIMP-2". J. Biol. Chem. 272 (24): 15496–500. doi:10.1074/jbc.272.24.15496. PMID 9182583.
- Kai HS, Butler GS, Morrison CJ, King AE, Pelman GR, Overall CM (December 2002). "Utilization of a novel recombinant myoglobin fusion protein expression system to characterize the tissue inhibitor of metalloproteinase (TIMP)-4 and TIMP-2 C-terminal domain and tails by mutagenesis. The importance of acidic residues in binding the MMP-2 hemopexin C-domain". J. Biol. Chem. 277 (50): 48696–707. doi:10.1074/jbc.M209177200. PMID 12374789.
Further reading
- Massova I, Kotra LP, Fridman R, Mobashery S (1998). "Matrix metalloproteinases: structures, evolution, and diversification". FASEB J. 12 (25n26): 1075–95. CiteSeerX 10.1.1.31.3959. doi:10.1142/S0217984998001256. PMID 9737711.
- Nagase H, Woessner JF (1999). "Matrix metalloproteinases". J. Biol. Chem. 274 (31): 21491–4. doi:10.1074/jbc.274.31.21491. PMID 10419448.
- Goffin F, Frankenne F, Béliard A, Perrier D'Hauterive S, Pignon MR, Geenen V, Foidart JM (2002). "Human endometrial epithelial cells modulate the activation of gelatinase a by stromal cells". Gynecol. Obstet. Invest. 53 (2): 105–11. doi:10.1159/000053003. PMID 11961384. S2CID 45390394.
- Hrabec E, Naduk J, Strek M, Hrabec Z (2007). "[Type IV collagenases (MMP-2 and MMP-9) and their substrates--intracellular proteins, hormones, cytokines, chemokines and their receptors]". Postepy Biochem. 53 (1): 37–45. PMID 17718386.
External links
PDB gallery | |
|---|---|
|