Haplogroup T (mtDNA)
Haplogroup T is a human mitochondrial DNA (mtDNA) haplogroup. It is believed to have originated around 25,100 years ago in the Near East.
| Haplogroup T | |
|---|---|
| Possible time of origin | 25,149 ± 4,668 years before present |
| Possible place of origin | Near East |
| Ancestor | JT |
| Descendants | T1 and T2 |
| Defining mutations | G709A, G1888A, A4917G, G8697A, T10463C, G13368A, G14905A, A15607G, G15928A, C16294T |
Origins
Mitochondrial clade T derives from the haplogroup JT, which also gave rise to the mtDNA haplogroup J. The T maternal clade is thought to have emanated from the Near East (Bermisheva 2002).
Distribution
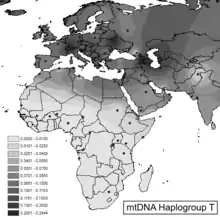
The basal haplogroup T* is found among Algerians in Oran (1.67%) and Reguibate Sahrawi (0.93%).[1] It is also distributed among the Soqotri (1.2%).[2]
Haplogroup T is present at low frequencies haplogroup throughout Western and Central Asia and Europe, with varying degrees of prevalence and certainly might have been present in other groups from the surrounding areas. T is found in approximately 10% of native Europeans.[3][4] It is also common among modern day Iranians. Based on a sample of over 400 modern day Iranians (Kivisild and Metspalu 2003), the T haplogroup represents roughly 8.3% of the population (about 1 out of 12 individuals), with the more specific T1 subtype constituting roughly half of those. Furthermore, the specific subtype T1 tends to be found further east and is common in Central Asian and modern Turkic populations (Lalueza-Fox 2004), who inhabit much of the same territory as the ancient Saka, Sarmatian, Andronovo, and other putative Iranian peoples of the 2nd and 1st millennia BC. Lalueza-Fox et al. (2004) also found several T and T1 sequences in ancient burials, including Kurgans, in the Kazakh steppe between the 14th-10th centuries BC, as well as later into the 1st millennia BC. These coincide with the latter part of the Andronovo period and the Saka period in the region.[5]
The geographic distribution within subclade T2 varies greatly with the ratio of subhaplogroup T2e to T2b reported to vary 40-fold across examined populations from a low in Britain and Ireland, to a high in Saudi Arabia (Bedford 2012). Within subhaplogroup T2e, a very rare motif is identified among Sephardic Jews of Turkey and Bulgaria and suspected conversos from the New World (Bedford 2012).
Found in Svan population from Caucasus (Georgia) T* 10,4% and T1 4,2%. T1a1a1 is particularly common in countries with high levels of Y-haplogroup R1a, such as Central and Northeast Europe. The clade is also found everywhere in Central Asia and deep into North Asia, as far east as Mongolia.
T2c and T2d appear to have a Near Eastern origin around the time of the Last Glacial Maximum (LGM) and more recent dispersals into Europe. Most of T2c comprises haplogroup T2c1. Apart from a peak in Cyprus, T2c1 is most common in the Persian Gulf region but is also found in the Levant and in Mediterranean Europe, with a more far-flung distribution at very low levels.[6]
T2 is also found among the Soqotri (7.7%).[2]
Archaeology
Wilde et al. (2014) tested mtDNA samples from the Yamna culture, the presumed homeland of Proto-Indo-European speakers. They found T2a1b in the Middle Volga region and Bulgaria, and T1a both in central Ukraine and the Middle Volga. The frequency of T1a and T2 in Yamna samples were each 14.5%, a percentage higher than in any country today and only found in similarly high frequencies among the Udmurts of the Volga-Ural region.[7]
Haplogroup T has also been found among Iberomaurusian specimens dating from the Epipaleolithic at the Afalou prehistoric site in Algeria. One ancient individual carried the T2b subclade (1/9; 11%).[8] Additionally, haplogroup T has been observed among ancient Egyptian mummies excavated at the Abusir el-Meleq archaeological site in Middle Egypt, which date from the Pre-Ptolemaic/late New Kingdom (T1, T2), Ptolemaic (T1, T2), and Roman (undifferentiated T, T1) periods.[9] Fossils excavated at the Late Neolithic site of Kelif el Boroud in Morocco, which have been dated to around 3,000 BCE, have also been observed to carry the T2 subclade.[10] Additionally, haplogroup T has been observed in ancient Guanche fossils excavated in Gran Canaria and Tenerife on the Canary Islands, which have been radiocarbon-dated to between the 7th and 11th centuries CE. The clade-bearing individuals were inhumed at the Tenerife site, with one specimen found to belong to the T2c1d2 subclade (1/7; 14%).[11]
Africa
In Africa, haplogroup T is primarily found among Afro-Asiatic-speaking populations, including the basal T* clade.[1] Some non-basal T clades are also commonly found among the Niger-Congo-speaking Serer due to diffusion from the Maghreb, likely with the spread of Islam and urban civilizations.[12]
| Population | Location | Language Family | N | Frequency | Source |
|---|---|---|---|---|---|
| Amhara | Ethiopia | Afro-Asiatic > Semitic | 5/120 | 4.17% | Kivisild 2004 |
| Beja | Sudan | Afro-Asiatic > Cushitic | 1/48 | 2.1% | Hassan 2009 |
| Beta Israel | Ethiopia | Afro-Asiatic > Cushitic | 0/29 | 0.00% | Behar 2008a |
| Copt | Egypt | Afro-Asiatic > Egyptian | 5/29 | 17.2% | Hassan 2009 |
| Dawro K. | Ethiopia | Afro-Asiatic > Omotic | 2/137 | 1.46% | Castrì 2008 and Boattini 2013 |
| Egyptians (El-Hayez) | Egypt | Afro-Asiatic > Semitic | 10/35 | 28.6% | Kujanova 2009 |
| Ethiopia | Ethiopia | Undetermined | 2/77 | 2.60% | Soares 2011 |
| Ethiopian Jew | Ethiopia | Afro-Asiatic > Cushitic | 0/41 | 0.00% | Non 2011 |
| Gurage | Ethiopia | Afro-Asiatic > Semitic | 0/21 | 0.00% | Kivisild 2004 |
| Hamer | Ethiopia | Afro-Asiatic > Omotic | 0/11 | 0.00% | Castrì 2008 and Boattini 2013 |
| Ongota | Ethiopia | Afro-Asiatic > Cushitic | 0/19 | 0.00% | Castrì 2008 and Boattini 2013 |
| Oromo | Ethiopia | Afro-Asiatic > Cushitic | 0/33 | 0.00% | Kivisild 2004 |
| Tigrai | Ethiopia | Afro-Asiatic > Semitic | 3/44 | 6.82% | Kivisild 2004 |
| Daasanach | Kenya | Afro-Asiatic > Cushitic | 0/49 | 0.00% | Poloni 2009 |
| Elmolo | Kenya | Afro-Asiatic > Cushitic | 0/52 | 0.00% | Castrì 2008 and Boattini 2013 |
| Luo | Kenya | Nilo-Saharan | 0/49 | 0.00% | Castrì 2008 and Boattini 2013 |
| Maasai | Kenya | Nilo-Saharan | 0/81 | 0.00% | Castrì 2008 and Boattini 2013 |
| Nairobi | Kenya | Niger-Congo | 0/100 | 0.00% | Brandstatter 2004 |
| Nyangatom | Kenya | Nilo-Saharan | 0/112 | 0.00% | Poloni 2009 |
| Rendille | Kenya | Afro-Asiatic > Cushitic | 0/17 | 0.00% | Castrì 2008 and Boattini 2013 |
| Samburu | Kenya | Nilo-Saharan | 0/35 | 0.00% | Castrì 2008 and Boattini 2013 |
| Turkana | Kenya | Nilo-Saharan | 0/51 | 0.00% | Castrì 2008 and Boattini 2013 |
| Hutu | Rwanda | Niger-Congo | 0/42 | 0.00% | Castrì 2009 |
| Dinka | Sudan | Nilo-Saharan | 0/46 | 0.00% | Krings 1999 |
| Sudan | Sudan | Undetermined | 3/102 | 2.94% | Soares 2011 |
| Burunge | Tanzania | Afro-Asiatic > Cushitic | 0/38 | 0.00% | Tishkoff 2007 |
| Datoga | Tanzania | Nilo-Saharan | 1/57 | 1.75% | Tishkoff 2007 and Knight 2003 |
| Iraqw | Tanzania | Afro-Asiatic > Cushitic | 0/12 | 0.00% | Knight 2003 |
| Sukuma | Tanzania | Niger-Congo | 0/32 | 0.00% | Tishkoff 2007 and Knight 2003 |
| Turu | Tanzania | Niger-Congo | 0/29 | 0.00% | Tishkoff 2007 |
| Yemeni | Yemen | Afro-Asiatic > Semitic | 1/114 | 0.88% | Kivisild 2004 |
Subclades
Tree
This phylogenetic tree of haplogroup I subclades is based on the paper (van Oven 2008) and subsequent published research (Behar 2012b). For brevity, only the first three levels of subclades (branches) are shown.
- T
- T1
- T1a
- T1a1
- T1b
- T1a
- T2
- T2a
- T2a1
- T2b
- T2b1
- T2b2
- T2b3
- T2b4
- T2b5
- T2b6
- T2c
- T2c1
- T2d
- T2e
- T2e2
- T2f
- T2f1
- T2g
- T2a
- T1
Health issues
One study has shown Haplogroup T to be associated with increased risk for coronary artery disease (Sanger 2007). However, some studies have also shown that people of Haplogroup T are less prone to diabetes (Chinnery 2007 and González 2012).
A few tentative medical studies have demonstrated that Haplogroup T may offer some resistance to both Parkinson's disease and Alzheimer's disease.
One study has found that among the Spanish population, Hypertrophic CardioMyopathy (HCM) also referred to as Hypertrophic Obstructive CardioMyopathy or HOCM is more likely to happen in those of T2 ancestry than those in other maternal haplogroups.[13] It is unknown whether or not this is specific to this subclaude of haplogroup T or is a risk factor shared by all of haplogroup T. With a statistically significant difference found in such a small sample, it may be advisable for those of known haplogroup T maternal ancestry to be aware of this and have their physician check for evidence of this condition when having a routine exam at an early age. It is usually symptom-less and increases the risk of sudden cardiac death, which often happens to those of as early in life as teenagers and may affect those who are active and have no other risk factors.[14]
Certain medical studies had shown mitochondrial Haplogroup T to be associated with reduced sperm motility in males, although these results have been challenged (Mishmar 2002). According to the Departamento de Bioquimica y Biologica Molecular y Celular, Universidad de Zaragoza, Haplogroup T can predispose to asthenozoospermia (Ruiz-Pesini 2000). However, these findings have been disputed due to a small sample size in the study (Mishmar 2002).
Famous members
During the BBC One documentary Meet the Izzards, the actor and comedian Eddie Izzard learns that her mitochondrial DNA is of Haplogroup T, specifically the subclade T2f1a1.[15]
Nicholas II of Russia
The last Russian Tsar, Nicholas II, has been shown to be of Haplogroup T, specifically subclade T2 (Ivanov 1996). Assuming all relevant pedigrees are correct, this includes all female-line descendants of his female line ancestor Barbara of Celje (1390–1451), wife of Sigismund, Holy Roman Emperor. This includes a great number of European nobles, including George I of Great Britain and Frederick William I of Prussia (through the Electress Sophia of Hanover), Charles I of England, George III of the United Kingdom, George V of the United Kingdom, Charles X Gustav of Sweden, Gustavus Adolphus of Sweden, Maurice of Nassau, Prince of Orange, Olav V of Norway, and George I of Greece.
References
Footnotes
Citations
- Asmahan Bekada; Lara R. Arauna; Tahria Deba; Francesc Calafell; Soraya Benhamamouch; David Comas (September 24, 2015). "Genetic Heterogeneity in Algerian Human Populations". PLOS ONE. 10 (9): e0138453. Bibcode:2015PLoSO..1038453B. doi:10.1371/journal.pone.0138453. PMC 4581715. PMID 26402429.; S5 Table
- Černý, Viktor; et al. (2009). "Out of Arabia—the settlement of island Soqotra as revealed by mitochondrial and Y chromosome genetic diversity" (PDF). American Journal of Physical Anthropology. 138 (4): 439–447. doi:10.1002/ajpa.20960. PMID 19012329. Archived from the original (PDF) on 6 October 2016. Retrieved 13 June 2016.
- Bryan Sykes (2001). The Seven Daughters of Eve. London; New York: Bantam Press. ISBN 978-0393020182.
- "Maternal Ancestry". Oxford Ancestors. Archived from the original on 15 July 2017. Retrieved 7 February 2013.
- Bennett, Casey; Kaestle, Frederika A. (2010). "Investigation of Ancient DNA from Western Siberia and the Sargat Culture". Human Biology. 82 (2): 143–156. arXiv:1112.2014. doi:10.3378/027.082.0202. PMID 20649397. S2CID 54566651.
- Pala, M; Olivieri, A; Achilli, A; Accetturo, M; Metspalu, E; Reidla, M; Tamm, E; Karmin, M; Reisberg, T; Hooshiar Kashani, B; Perego, UA; Carossa, V; Gandini, F; Pereira, JB; Soares, P; Angerhofer, N; Rychkov, S; Al-Zahery, N; Carelli, V; Sanati, MH; Houshmand, M; Hatina, J; Macaulay, V; Pereira, L; Woodward, SR; Davies, W; Gamble, C; Baird, D; Semino, O; Villems, R; Torroni, A; Richards, MB (4 May 2012). "Mitochondrial DNA Signals of Late Glacial Recolonization of Europe from Near Eastern Refugia". The American Journal of Human Genetics. 90 (5): 915–924. doi:10.1016/j.ajhg.2012.04.003. PMC 3376494. PMID 22560092.http://haplogroup.org/sources/mitochondrial-dna-signals-of-late-glacial-recolonization-of-europe-from-near-eastern-refugia/
- Wilde, Sandra (2014). "Direct evidence for positive selection of skin, hair, and eye pigmentation in Europeans during the last 5,000 y". Proceedings of the National Academy of Sciences. 111 (13): 4832–4837. Bibcode:2014PNAS..111.4832W. doi:10.1073/pnas.1316513111. PMC 3977302. PMID 24616518.
- Kefi, Rym; et al. (2018). "On the origin of Iberomaurusians: new data based on ancient mitochondrial DNA and phylogenetic analysis of Afalou and Taforalt populations". Mitochondrial DNA Part A. 29 (1): 147–157. doi:10.1080/24701394.2016.1258406. PMID 28034339. S2CID 4490910.
- Schuenemann, Verena J.; et al. (2017). "Ancient Egyptian mummy genomes suggest an increase of Sub-Saharan African ancestry in post-Roman periods". Nature Communications. 8: 15694. Bibcode:2017NatCo...815694S. doi:10.1038/ncomms15694. PMC 5459999. PMID 28556824.
- Fregel; et al. (2018). "Ancient genomes from North Africa evidence prehistoric migrations to the Maghreb from both the Levant and Europe" (PDF). bioRxiv 10.1101/191569.
- Rodrı́guez-Varela; et al. (2017). "Genomic Analyses of Pre-European Conquest Human Remains from the Canary Islands Reveal Close Affinity to Modern North Africans". Current Biology. 27 (1–7): 3396–3402.e5. doi:10.1016/j.cub.2017.09.059. PMID 29107554. Retrieved 28 October 2017.
- Ball, Edward (2007). The Genetic Strand: Exploring a Family History Through DNA. Simon and Schuster. p. 233. ISBN 978-1416554257. Retrieved 31 May 2016.
- Castro, M (2006). "Mitochondrial DNA haplogroups in Spanish patients with hypertrophic cardiomyopathy". Int J Cardiol. 112 (2): 202–6. doi:10.1016/j.ijcard.2005.09.008. PMID 16313983.
- Chen, Michael. "Hypertrophic cardiomyopathy - Medical Encyclopedia". Medline Plus. National Library of Medicine. Retrieved 2015-10-03.
- Meet the Izzards: The Mum's Line. BBC One. 2013-03-12. 48 minutes in.
- Pike DA, Barton TJ, Bauer SL, Kipp E (2010). "mtDNA Haplogroup T Phylogeny Based on Full Mitochondrial Sequences". Journal of Genetic Genealogy. 6 (1).
- "Molecular instability of the mitochondrial haplogroup T sequences at nucleotide positions 16292 and 16296". Cite journal requires
|journal=(help) - Abu-Amero, Khaled K; Larruga, José M; Cabrera, Vicente M; González, Ana M (2008). "Mitochondrial DNA structure in the Arabian Peninsula". BMC Evolutionary Biology. 8: 45. doi:10.1186/1471-2148-8-45. PMC 2268671. PMID 18269758.
- Babalini, Carla; Martínez-Labarga, Cristina; Tolk, Helle-Viivi; Kivisild, Toomas; Giampaolo, Rita; Tarsi, Tiziana; Contini, Irene; Barać, Lovorka; et al. (2005). "The population history of the Croatian linguistic minority of Molise (southern Italy): A maternal view". European Journal of Human Genetics. 13 (8): 902–12. doi:10.1038/sj.ejhg.5201439. PMID 15886710.
- Bedford, Felice L (2012). "Sephardic signature in haplogroup T mitochondrial DNA". European Journal of Human Genetics. 20 (4): 441–8. doi:10.1038/ejhg.2011.200. PMC 3306851. PMID 22108605.
- Behar, DM; Metspalu, E; Kivisild, T; Rosset, S; Tzur, S; Hadid, Y; Yudkovsky, G; Rosengarten, D; et al. (2008). MacAulay, Vincent (ed.). "Counting the founders: The matrilineal genetic ancestry of the Jewish Diaspora". PLOS ONE. 3 (4): e2062. Bibcode:2008PLoSO...3.2062B. doi:10.1371/journal.pone.0002062. PMC 2323359. PMID 18446216.
- Behar, Doron M.; Van Oven, Mannis; Rosset, Saharon; Metspalu, Mait; Loogväli, Eva-Liis; Silva, Nuno M.; Kivisild, Toomas; Torroni, Antonio; Villems, Richard (2012). "A "Copernican" Reassessment of the Human Mitochondrial DNA Tree from its Root". The American Journal of Human Genetics. 90 (4): 675–84. doi:10.1016/j.ajhg.2012.03.002. PMC 3322232. PMID 22482806.
- Belyaeva, Olga; Bermisheva, Marina; Khrunin, Andrey; Slominsky, Petr; Bebyakova, Natalia; Khusnutdinova, E. K. (Elza Kamilevna); Mikulich, Aleksei Ignatevich; Limborskaia, S. A. (Svetlana Andreevna) (2003). "Mitochondrial DNA variations in Russian and Belorussian populations". Human Biology. 75 (5): 647–60. doi:10.1353/hub.2003.0069. PMID 14763602. S2CID 23876546.
- Boattini, Alessio; Castrì, Loredana; Sarno, Stefania; Useli, Antonella; Cioffi, Manuela; Sazzini, Marco; Garagnani, Paolo; De Fanti, Sara; Pettener, Davide; Luiselli, Donata (2013). "MtDNA variation in East Africa unravels the history of afro-asiatic groups". American Journal of Physical Anthropology. 150 (3): 375–385. doi:10.1002/ajpa.22212. PMID 23283748.
- Bosch, E.; Calafell, F.; Gonzalez-Neira, A.; Flaiz, C.; Mateu, E.; Scheil, H.-G.; Huckenbeck, W.; Efremovska, L.; et al. (2006). "Paternal and maternal lineages in the Balkans show a homogeneous landscape over linguistic barriers, except for the isolated Aromuns". Annals of Human Genetics. 70 (4): 459–87. doi:10.1111/j.1469-1809.2005.00251.x. PMID 16759179. S2CID 23156886.
- Brandstatter, Anita; Peterson, Christine T.; Irwin, Jodi A.; Mpoke, Solomon; Koech, Davy K.; Parson, Walther; Parsons, Thomas J. (2004). "Mitochondrial DNA control region sequences from Nairobi (Kenya): Inferring phylogenetic parameters for the establishment of a forensic database". International Journal of Legal Medicine. 118 (5): 294–306. doi:10.1007/s00414-004-0466-z. PMID 15248073. S2CID 19703169.
- Castrì, Loredana; Garagnani, P; Useli, A; Pettener, D; Luiselli, D (2008). "Kenyan crossroads: migration and gene flow in six ethnic groups from Eastern Africa". Journal of Anthropological Sciences. 86: 189–192. PMID 19934476.
- Castrì, Loredana; Tofanelli, Sergio; Garagnani, Paolo; Bini, Carla; Fosella, Xenia; Pelotti, Susi; Paoli, Giorgio; Pettener, Davide; Luiselli, Donata (2009). "MtDNA variability in two Bantu-speaking populations (Shona and Hutu) from Eastern Africa: Implications for peopling and migration patterns in sub-Saharan Africa". American Journal of Physical Anthropology. 140 (2): 302–11. doi:10.1002/ajpa.21070. PMID 19425093.
- Chinnery, P F; Mowbray, C; Patel, S K; Elson, J L; Sampson, M; Hitman, G A; McCarthy, M I; Hattersley, A T; Walker, M (2007). "Mitochondrial DNA haplogroups and type 2 diabetes: A study of 897 cases and 1010 controls". Journal of Medical Genetics. 44 (6): e80. doi:10.1136/jmg.2007.048876. PMC 2740896. PMID 17551080.
- Coble, Michael D.; Loreille, Odile M.; Wadhams, Mark J.; Edson, Suni M.; Maynard, Kerry; Meyer, Carna E.; Niederstätter, Harald; Berger, Cordula; et al. (2009). Hofreiter, Michael (ed.). "Mystery Solved: The Identification of the Two Missing Romanov Children Using DNA Analysis". PLOS ONE. 4 (3): e4838. Bibcode:2009PLoSO...4.4838C. doi:10.1371/journal.pone.0004838. PMC 2652717. PMID 19277206.
- Costa, MD; Cherni, L; Fernandes, V; Freitas, F; Ammar El Gaaied, AB; Pereira, L (2009). "Data from complete mtDNA sequencing of Tunisian centenarians: Testing haplogroup association and the "golden mean" to longevity". Mechanisms of Ageing and Development. 130 (4): 222–6. doi:10.1016/j.mad.2008.12.001. PMID 19133286. S2CID 6102820.
- Cvjetan, S; Tolk, HV; Lauc, LB; Colak, I; Dordević, D; Efremovska, L; Janićijević, B; Kvesić, A; et al. (2004). "Frequencies of mtDNA haplogroups in southeastern Europe--Croatians, Bosnians and Herzegovinians, Serbians, Macedonians and Macedonian Romani". Collegium Antropologicum. 28 (1): 193–8. PMID 15636075.
- Derenko, M; Malyarchuk, B; Grzybowski, T; Denisova, G; Dambueva, I; Perkova, M; Dorzhu, C; Luzina, F; et al. (2007). "Phylogeographic analysis of mitochondrial DNA in northern Asian populations". American Journal of Human Genetics. 81 (5): 1025–41. doi:10.1086/522933. PMC 2265662. PMID 17924343.
- Dubut, Vincent; Chollet, Lionel; Murail, Pascal; Cartault, François; Béraud-Colomb, Eliane; Serre, Myriam; Mogentale-Profizi, Nérina (2003). "MtDNA polymorphisms in five French groups: Importance of regional sampling". European Journal of Human Genetics. 12 (4): 293–300. doi:10.1038/sj.ejhg.5201145. PMID 14694359.
- Fernandes, Verónica; Alshamali, Farida; Alves, Marco; Costa, Marta D.; Pereira, Joana B.; Silva, Nuno M.; Cherni, Lotfi; Harich, Nourdin; et al. (2012). "The Arabian Cradle: Mitochondrial Relicts of the First Steps along the Southern Route out of Africa". The American Journal of Human Genetics. 90 (2): 347–355. doi:10.1016/j.ajhg.2011.12.010. PMC 3276663. PMID 22284828.
- Finnila, JS; Finnila, S; Majamaa, K (2001). "Lineage-specific selection in human mtDNA: Lack of polymorphisms in a segment of MTND5 gene in haplogroup J". Molecular Biology and Evolution. 20 (12): 2132–42. doi:10.1093/molbev/msg230. PMID 12949126.
- Gasparre, G.; Porcelli, A. M.; Bonora, E.; Pennisi, L. F.; Toller, M.; Iommarini, L.; Ghelli, A.; Moretti, M.; et al. (2007). "Disruptive mitochondrial DNA mutations in complex I subunits are markers of oncocytic phenotype in thyroid tumors". Proceedings of the National Academy of Sciences. 104 (21): 9001–9006. Bibcode:2007PNAS..104.9001G. doi:10.1073/pnas.0703056104. PMC 1885617. PMID 17517629.
- Gonder, M. K.; Mortensen, H. M.; Reed, F. A.; De Sousa, A.; Tishkoff, S. A. (2006). "Whole-mtDNA Genome Sequence Analysis of Ancient African Lineages". Molecular Biology and Evolution. 24 (3): 757–68. doi:10.1093/molbev/msl209. PMID 17194802.
- González, Ana M.; MacEira, Benito M.; Pérez, Estefanía; Cabrera, Vicente M.; López, Alfonso J.; Larruga, José M. (2012). "Genetics, Environment, and Diabetes-Related End-Stage Renal Disease in the Canary Islands". Genetic Testing and Molecular Biomarkers. 16 (8): 859–64. doi:10.1089/gtmb.2011.0207. PMC 3422557. PMID 22480375.
- Hartmann, A; Thieme, M; Nanduri, LK; Stempfl, T; Moehle, C; Kivisild, T; Oefner, PJ (2009). "Validation of microarray-based resequencing of 93 worldwide mitochondrial genomes". Human Mutation. 30 (1): 115–22. doi:10.1002/humu.20816. PMID 18623076. S2CID 205918494.
- Hassan, Hisham Yousif (2009). "Genetic Patterns of Y-chromosome and Mitochondrial DNA Variation, with Implications to the Peopling of the Sudan" (PDF). University of Khartoum.
- Helgason, Agnar; Hickey, Eileen; Goodacre, Sara; Bosnes, Vidar; Stefánsson, Kári; Ward, Ryk; Sykes, Bryan (2001). "mtDNA and the Islands of the North Atlantic: Estimating the Proportions of Norse and Gaelic Ancestry". The American Journal of Human Genetics. 68 (3): 206–15. doi:10.1086/318785. PMC 1274484. PMID 11179019.
- Hofreiter, Linea; Lynnerup, Niels; Siegismund, Hans R.; Kivisild, Toomas; Dissing, Jørgen (2010). Hofreiter, Michael (ed.). "Genetic Diversity among Ancient Nordic Populations". PLOS ONE. 5 (7): e11898. Bibcode:2010PLoSO...511898M. doi:10.1371/journal.pone.0011898. PMC 2912848. PMID 20689597.
The overall occurrence of haplogroups did not deviate from extant Scandinavians, however, haplogroup I was significantly more frequent among the ancient Danes (average 13%) than among extant Danes and Scandinavians (~2.5%) as well as among other ancient population samples reported. Haplogroup I could therefore have been an ancient Southern Scandinavian type “diluted” by later immigration events.
- Ivanov, Pavel L.; Wadhams, Mark J.; Roby, Rhonda K.; Holland, Mitchell M.; Weedn, Victor W.; Parsons, Thomas J. (1996). "Mitochondrial DNA sequence heteroplasmy in the Grand Duke of Russia Georgij Romanov establishes the authenticity of the remains of Tsar Nicholas II". Nature Genetics. 12 (4): 417–20. doi:10.1038/ng0496-417. PMID 8630496. S2CID 287478.
- Janssen, GM; Neu, A; 't Hart, LM; Van De Sande, CM; Antonie Maassen, J (2006). "Novel mitochondrial DNA length variants and genetic instability in a family with diabetes and deafness". Experimental and Clinical Endocrinology & Diabetes. 114 (4): 168–74. doi:10.1055/s-2006-924066. PMID 16705548.
- Keyser, Christine; Bouakaze, Caroline; Crubézy, Eric; Nikolaev, Valery G.; Montagnon, Daniel; Reis, Tatiana; Ludes, Bertrand (2009). "Ancient DNA provides new insights into the history of south Siberian Kurgan people". Human Genetics. 126 (3): 395–410. doi:10.1007/s00439-009-0683-0. PMID 19449030. S2CID 21347353.
- Kivisild, T; Reidla, M; Metspalu, E; Rosa, A; Brehm, A; Pennarun, E; Parik, J; Geberhiwot, T; et al. (2004). "Ethiopian mitochondrial DNA heritage: Tracking gene flow across and around the gate of tears". American Journal of Human Genetics. 75 (5): 752–70. doi:10.1086/425161. PMC 1182106. PMID 15457403.
- Knight, A; Underhill, PA; Mortensen, HM; Zhivotovsky, LA; Lin, AA; Henn, BM; Louis, D; Ruhlen, M; Mountain, JL (2003). "African Y chromosome and mtDNA divergence provides insight into the history of click languages". Current Biology. 13 (6): 464–73. doi:10.1016/S0960-9822(03)00130-1. PMID 12646128. S2CID 52862939.
- Krings, M; Salem, AE; Bauer, K; Geisert, H; Malek, AK; Chaix, L; Simon, C; Welsby, D; et al. (1999). "MtDNA analysis of Nile River Valley populations: A genetic corridor or a barrier to migration?". American Journal of Human Genetics. 64 (4): 1166–1176. doi:10.1086/302314. PMC 1377841. PMID 10090902.
- Martina Kujanova; Luisa Pereira; Veronica Fernandes; Joana B. Pereira; Viktor Cerny (2009). "Near Eastern Neolithic Genetic Input in a Small Oasis of the Egyptian Western Desert". American Journal of Physical Anthropology. 140 (2): 336–346. doi:10.1002/ajpa.21078. PMID 19425100.
- Lalueza-Fox, C; Sampietro, ML; Gilbert, MT; Castri, L; Facchini, F; Pettener, D; Bertranpetit, J (2004). "Unravelling migrations in the steppe: Mitochondrial DNA sequences from ancient central Asians". Proceedings: Biological Sciences. 271 (1542): 941–7. doi:10.1098/rspb.2004.2698. PMC 1691686. PMID 15255049.
- Maca-Meyer, N; González, AM; Larruga, JM; Flores, C; Cabrera, VM (2001). "Major genomic mitochondrial lineages delineate early human expansions". BMC Genetics. 2: 13. doi:10.1186/1471-2156-2-13. PMC 55343. PMID 11553319.
- MacAulay, Vincent; Richards, Martin; Hickey, Eileen; Vega, Emilce; Cruciani, Fulvio; Guida, Valentina; Scozzari, Rosaria; Bonné-Tamir, Batsheva; et al. (1999). "The Emerging Tree of West Eurasian mtDNAs: A Synthesis of Control-Region Sequences and RFLPs". The American Journal of Human Genetics. 64 (1): 232–49. doi:10.1086/302204. PMC 1377722. PMID 9915963.
- Malyarchuk, B. A.; Derenko, M. V. (1999). "Molecular instability of the mitochondrial haplogroup T sequences at nucleotide positions 16292 and 16296". Annals of Human Genetics. 63 (6): 489–497. doi:10.1046/j.1469-1809.1999.6360489.x. PMID 11246451. S2CID 45007072.
- Malyarchuk, B. A.; Derenko, M. V. (2001). "Mitochondrial DNA variability in Russians and Ukrainians: Implication to the origin of the Eastern Slavs". Annals of Human Genetics. 65 (Pt 1): 63–78. doi:10.1046/j.1469-1809.2001.6510063.x. PMID 11415523. S2CID 9392520.
- Malyarchuk, BA; Grzybowski, T; Derenko, MV; Czarny, J; Drobnic, K; Miścicka-Sliwka, D (2003). "Mitochondrial DNA variability in Bosnians and Slovenians". Annals of Human Genetics. 67 (Pt 5): 412–25. doi:10.1046/j.1469-1809.2003.00042.x. PMID 12940915. S2CID 2105448.
- Malyarchuk, B; Derenko, M; Denisova, G; Kravtsova, O (2010). "Mitogenomic diversity in Tatars from the Volga-Ural region of Russia". Molecular Biology and Evolution. 27 (10): 2220–6. doi:10.1093/molbev/msq065. PMID 20457583.
- Martinez-Cruz, B.; Harmant, C.; Platt, D. E.; Haak, W.; Manry, J.; Ramos-Luis, E.; Soria-Hernanz, D. F.; Bauduer, F.; et al. (2012). "Evidence of Pre-Roman Tribal Genetic Structure in Basques from Uniparentally Inherited Markers". Molecular Biology and Evolution. 29 (9): 2211–22. doi:10.1093/molbev/mss091. PMID 22411853.
- Melchior, Linea; Kivisild, Toomas; Lynnerup, Niels; Dissing, Jørgen (2008). Ahmed, Niyaz (ed.). "Evidence of Authentic DNA from Danish Viking Age Skeletons Untouched by Humans for 1,000 Years". PLOS ONE. 3 (5): e2214. Bibcode:2008PLoSO...3.2214M. doi:10.1371/journal.pone.0002214. PMC 2386972. PMID 18509537.
- Metspalu, Mait; Kivisild, Toomas; Metspalu, Ene; Parik, Jüri; Hudjashov, Georgi; Kaldma, Katrin; Serk, Piia; Karmin, Monika; Behar, Doron M; Gilbert, M Thomas P; Endicott, Phillip; Mastana, Sarabjit; Papiha, Surinder S; Skorecki, Karl; Torroni, Antonio; Villems, Richard (2004). "Most of the extant mtDNA boundaries in south and southwest Asia were likely shaped during the initial settlement of Eurasia by anatomically modern humans". BMC Genetics. 5: 26. doi:10.1186/1471-2156-5-26. PMC 516768. PMID 15339343.
- Mikkelsen, Martin; Sørensen, Erik; Rasmussen, Erik Michael; Morling, Niels (2010). "Mitochondrial DNA HV1 and HV2 variation in Danes". Forensic Science International: Genetics. 4 (4): e87–8. doi:10.1016/j.fsigen.2009.07.007. PMID 20457038.
- Mishmar, D; Ruiz-Pesini, E; Golik, P; MacAulay, V; Clark, AG; Hosseini, S; Brandon, M; Easley, K; et al. (2003). "Natural selection shaped regional mtDNA variation in humans". Proceedings of the National Academy of Sciences of the United States of America. 100 (1): 171–6. Bibcode:2003PNAS..100..171M. doi:10.1073/pnas.0136972100. PMC 140917. PMID 12509511.
- Musilová, Eliška; Fernandes, Verónica; Silva, Nuno M.; Soares, Pedro; Alshamali, Farida; Harich, Nourdin; Cherni, Lotfi; Gaaied, Amel Ben Ammar El; et al. (2011). "Population history of the Red Sea-genetic exchanges between the Arabian Peninsula and East Africa signaled in the mitochondrial DNA HV1 haplogroup". American Journal of Physical Anthropology. 145 (4): 592–8. doi:10.1002/ajpa.21522. PMID 21660931.
- Nikitin, Alexey G.; Kochkin, Igor T.; June, Cynthia M.; Willis, Catherine M.; McBain, Ian; Videiko, Mykhailo Y. (2009). "Mitochondrial DNA Sequence Variation in the Boyko, Hutsul, and Lemko Populations of the Carpathian Highlands". Human Biology. 81 (1): 43–58. doi:10.3378/027.081.0104. PMID 19589018. S2CID 45791162.
- Non, Amy L.; Al-Meeri, Ali; Raaum, Ryan L.; Sanchez, Luisa F.; Mulligan, Connie J. (2011). "Mitochondrial DNA reveals distinct evolutionary histories for Jewish populations in Yemen and Ethiopia". American Journal of Physical Anthropology. 144 (1): 1–10. doi:10.1002/ajpa.21360. PMID 20623605.
- Palanichamy, MG; Sun, C; Agrawal, S; Bandelt, HJ; Kong, QP; Khan, F; Wang, CY; Chaudhuri, TK; et al. (2004). "Phylogeny of mitochondrial DNA macrohaplogroup N in India, based on complete sequencing: Implications for the peopling of South Asia". American Journal of Human Genetics. 75 (6): 966–78. doi:10.1086/425871. PMC 1182158. PMID 15467980.
- Pereira, L; Gonçalves, J; Franco-Duarte, R; Silva, J; Rocha, T; Arnold, C; Richards, M; MacAulay, V (2007). "No evidence for an mtDNA role in sperm motility: Data from complete sequencing of asthenozoospermic males". Molecular Biology and Evolution. 24 (3): 868–74. doi:10.1093/molbev/msm004. PMID 17218641.
- Pericić, M; Barać Lauc, L; Martinović Klarić, I; Janićijević, B; Rudan, P (2005). "Review of Croatian genetic heritage as revealed by mitochondrial DNA and Y chromosomal lineages". Croatian Medical Journal. 46 (4): 502–13. PMID 16100752.
- Pichler, Irene; Fuchsberger, Christian; Platzer, Christa; Çalişkan, Minal; Marroni, Fabio; Pramstaller, Peter P; Ober, Carole (2010). "Drawing the history of the Hutterite population on a genetic landscape: Inference from Y-chromosome and mtDNA genotypes". European Journal of Human Genetics. 18 (4): 463–70. doi:10.1038/ejhg.2009.172. PMC 2987252. PMID 19844259.
- Poloni, Estella S.; Naciri, Yamama; Bucho, Rute; Niba, Régine; Kervaire, Barbara; Excoffier, Laurent; Langaney, André; Sanchez-Mazas, Alicia (2009). "Genetic Evidence for Complexity in Ethnic Differentiation and History in East Africa". Annals of Human Genetics. 73 (6): 582–600. doi:10.1111/j.1469-1809.2009.00541.x. PMID 19706029. S2CID 2488794.
- Pope, AM; Carr, SM; Smith, KN; Marshall, HD; Marshall, H. D. (2011). "Mitogenomic and microsatellite variation in descendants of the founder population of Newfoundland: High genetic diversity in an historically isolated population". Genome. 54 (2): 110–9. doi:10.1139/G10-102. PMID 21326367.
- Quintana-Murci, Lluís; Chaix, Raphaëlle; Wells, R. Spencer; Behar, Doron M.; Sayar, Hamid; Scozzari, Rosaria; Rengo, Chiara; Al-Zahery, Nadia; et al. (2004). "Where West Meets East: The Complex mtDNA Landscape of the Southwest and Central Asian Corridor". The American Journal of Human Genetics. 74 (5): 827–45. doi:10.1086/383236. PMC 1181978. PMID 15077202.
- Richards, Martin; MacAulay, Vincent; Hickey, Eileen; Vega, Emilce; Sykes, Bryan; Guida, Valentina; Rengo, Chiara; Sellitto, Daniele; et al. (2000). "Tracing European Founder Lineages in the Near Eastern mtDNA Pool". The American Journal of Human Genetics. 67 (5): 1251–76. doi:10.1016/S0002-9297(07)62954-1. PMC 1288566. PMID 11032788.
- Richards, Martin; Rengo, Chiara; Cruciani, Fulvio; Gratrix, Fiona; Wilson, James F.; Scozzari, Rosaria; MacAulay, Vincent; Torroni, Antonio (2003). "Extensive Female-Mediated Gene Flow from Sub-Saharan Africa into Near Eastern Arab Populations". The American Journal of Human Genetics. 72 (4): 1058–64. doi:10.1086/374384. PMC 1180338. PMID 12629598.
- Rogaev, E. I.; Grigorenko, A. P.; Moliaka, Y. K.; Faskhutdinova, G.; Goltsov, A.; Lahti, A.; Hildebrandt, C.; Kittler, E. L. W.; Morozova, I. (2009). "Genomic identification in the historical case of the Nicholas II royal family". Proceedings of the National Academy of Sciences. 106 (13): 5258–63. Bibcode:2009PNAS..106.5258R. doi:10.1073/pnas.0811190106. PMC 2664067. PMID 19251637.
- Ruiz-Pesini, Eduardo; Lapeña, Ana-Cristina; Díez-Sánchez, Carmen; Pérez-Martos, Acisclo; Montoya, Julio; Alvarez, Enrique; Díaz, Miguel; Urriés, Antonio; et al. (2000). "Human mtDNA Haplogroups Associated with High or Reduced Spermatozoa Motility". The American Journal of Human Genetics. 67 (3): 682–96. doi:10.1086/303040. PMC 1287528. PMID 10936107.
- Shlush, Liran I.; Behar, Doron M.; Yudkovsky, Guennady; Templeton, Alan; Hadid, Yarin; Basis, Fuad; Hammer, Michael; Itzkovitz, Shalev; Skorecki, Karl (2008). Gemmell, Neil John (ed.). "The Druze: A Population Genetic Refugium of the Near East". PLOS ONE. 3 (5): e2105. Bibcode:2008PLoSO...3.2105S. doi:10.1371/journal.pone.0002105. PMC 2324201. PMID 18461126.
- Soares, P.; Alshamali, F.; Pereira, J. B.; Fernandes, V.; Silva, N. M.; Afonso, C.; Costa, M. D.; Musilova, E.; et al. (2011). "The Expansion of mtDNA Haplogroup L3 within and out of Africa". Molecular Biology and Evolution. 29 (3): 915–27. doi:10.1093/molbev/msr245. PMID 22096215.
- Stanger, Olaf; Müller, Edith; Zimmermann, Franz; Wiesbauer, Martina; Mayr, Johannes A.; Paulweber, Bernhard; Iglseder, Bernhard; Renner, Wilfried; et al. (2007). "30 Mitochondrial haplogroup T is associated with coronary artery disease". Mitochondrion. 7 (6): 412. doi:10.1016/j.mito.2007.08.034.
- Stone, AC; Starrs, JE; Stoneking, M (2001). "Mitochondrial DNA analysis of the presumptive remains of Jesse James". Journal of Forensic Sciences. 46 (1): 173–6. doi:10.1520/JFS14932J. PMID 11210907.
- Terreros, Maria C; Rowold, Diane J; Mirabal, Sheyla; Herrera, Rene J (2011). "Mitochondrial DNA and Y-chromosomal stratification in Iran: Relationship between Iran and the Arabian Peninsula". Journal of Human Genetics. 56 (3): 235–46. doi:10.1038/jhg.2010.174. PMID 21326310.
- Thomas, Mark G; Barnes, Ian; Weale, Michael E; Jones, Abigail L; Forster, Peter; Bradman, Neil; Pramstaller, Peter P (2008). "New genetic evidence supports isolation and drift in the Ladin communities of the South Tyrolean Alps but not an ancient origin in the Middle East". European Journal of Human Genetics. 16 (1): 124–34. doi:10.1038/sj.ejhg.5201906. PMID 17712356.
- Tishkoff, S. A.; Gonder, M. K.; Henn, B. M.; Mortensen, H.; Knight, A.; Gignoux, C.; Fernandopulle, N.; Lema, G.; et al. (2007). "History of Click-Speaking Populations of Africa Inferred from mtDNA and Y Chromosome Genetic Variation". Molecular Biology and Evolution. 24 (10): 2180–95. doi:10.1093/molbev/msm155. PMID 17656633.
- Topf, A. L.; Gilbert, MT; Dumbacher, JP; Hoelzel, AR (2005). "Tracing the Phylogeography of Human Populations in Britain Based on 4th-11th Century mtDNA Genotypes". Molecular Biology and Evolution. 23 (1): 152–61. doi:10.1093/molbev/msj013. PMID 16151183.
- Torroni, A; Huoponen, K; Francalacci, P; Petrozzi, M; Morelli, L; Scozzari, R; Obinu, D; Savontaus, ML; Wallace, DC (1996). "Classification of European mtDNAs From an Analysis of Three European Populations". Genetics. 144 (4): 1835–50. PMC 1207732. PMID 8978068.
- van Oven, Mannis; Kayser, Manfred (2009). "Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation". Human Mutation. 30 (2): E386–94. doi:10.1002/humu.20921. PMID 18853457. S2CID 27566749.
Websites
- Behar; Family Tree DNA (2012). "mtDNA Community".
Further reading
- Černý, Viktor; Pereira, Luísa; Kujanová, Martina; VašÍková, Alžběta; Hájek, Martin; Morris, Miranda; Mulligan, Connie J. (2009). "Out of Arabia-The settlement of Island Soqotra as revealed by mitochondrial and Y chromosome genetic diversity". American Journal of Physical Anthropology. 138 (4): 439–47. doi:10.1002/ajpa.20960. PMID 19012329.
- Kitchen, A.; Ehret, C.; Assefa, S.; Mulligan, C. J. (2009). "Bayesian phylogenetic analysis of Semitic languages identifies an Early Bronze Age origin of Semitic in the Near East". Proceedings of the Royal Society B: Biological Sciences. 276 (1668): 2703–10. doi:10.1098/rspb.2009.0408. PMC 2839953. PMID 19403539.
- Petit-Maire, Nicole; Bouysse, Philippe (2000). "Geological records of the recent past, a key to the near future world environments" (PDF). Episodes. 23 (4): 230–246. doi:10.18814/epiiugs/2000/v23i4/001.
External links
- General
- Ian Logan's Mitochondrial DNA Site
- Mannis van Oven's Phylotree
- The Genographic Project Public Participation Mitochondrial DNA Database
- Haplogroup T
- Discussion List at RootsWeb
- Spread of Haplogroup T, from National Geographic
- Genetic Genealogy: A Personal Perspective on Tara, Karelians and Kent, England
- Analysis of a Haplogroup T sequence (T5/T2)
- Phylogenetic Networks for the Human mtDNA Haplogroup T
- mtDNA Haplogroup T - Full Genomic Sequence Research Project
- Phylogenetic Networks for the Human mtDNA Haplogroup T