Q10 (temperature coefficient)
The Q10 temperature coefficient is a measure of the rate of change of a biological or chemical system as a consequence of increasing the temperature by 10 °C. There are many examples where the Q10 is used, one being the calculation of the nerve conduction velocity and another being calculating the contraction velocity of muscle fibres. It can also be applied to chemical reactions and many other systems.

The Q10 is calculated as:

where
Rewriting this equation, the assumption behind Q10 is that the reaction rate R depends exponentially on temperature:

Q10 is a unitless quantity, as it is the factor by which a rate changes, and is a useful way to express the temperature dependence of a process.
For most biological systems, the Q10 value is ~ 2 to 3.[1]
In muscle performance
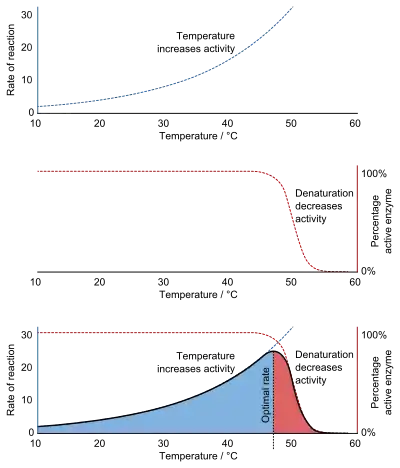
The temperature of a muscle has a significant effect on the velocity and power of the muscle contraction, with performance generally declining with decreasing temperatures and increasing with rising temperatures. The Q10 coefficient represents the degree of temperature dependence a muscle exhibits as measured by contraction rates. A Q10 of 1.0 indicates thermal independence of a muscle whereas an increasing Q10 value indicates increasing thermal dependence. Values less than 1.0 indicate a negative or inverse thermal dependence, i.e., a decrease in muscle performance as temperature increases.[2]
Q10 values for biological processes vary with temperature. Decreasing muscle temperature results in a substantial decline of muscle performance such that a 10 degree Celsius temperature decrease results in at least a 50% decline in muscle performance.[3] Persons who have fallen into icy water may gradually lose the ability to swim or grasp safety lines due to this effect, although other effects such as atrial fibrillation are a more immediate cause of drowning deaths. At some minimum temperature biological systems do not function at all, but performance increases with rising temperature (Q10 of 2-4) to a maximum performance level and thermal independence (Q10 of 1.0-1.5). With continued increase in temperature, performance decreases rapidly (Q10 of 0.2-0.8) up to a maximum temperature at which all biological function again ceases.[4]
Within vertebrates, different skeletal muscle activity has correspondingly different thermal dependencies. The rate of muscle twitch contractions and relaxations are thermally dependent (Q10 of 2.0-2.5), whereas maximum contraction, e.g., tetanic contraction, is thermally independent.[5]
Muscles of some ectothermic species. e.g., sharks, show less thermal dependence at lower temperatures than endothermic species [3][6]
See also
- Arrhenius equation
- Arrhenius plot
- Isotonic (exercise physiology)
- Isometric exercise
- Skeletal striated muscle
- Tetanic contraction
References
- Reyes, A.B.; Pendergast, J.S.; Yamazaki, S. (2008). "Mammalian peripheral circadian oscillators are temperature compensated". J. Biol. Rhythms. 23: 95–98. doi:10.1177/0748730407311855. PMC 2365757. PMID 18258762.
- Bennett, A.F. (1984). "Thermal dependence of muscle function". Am. J. Physiol. 247: R217–R229. PMID 6380314.
- Deban, Stephen M.; Lappin, A. Kristopher (2011). "Thermal effects on the dynamics and motor control of ballistic prey capture in toads: maintaining high performance at low temperature". The Journal of Experimental Biology. 214 (8): 1333–1346. doi:10.1242/jeb.048405. PMID 21430211.
- Bennett, A.F. (1990). "Thermal dependence of locomotor capacity". Am. J. Physiol. 259 (2 Pt 2): R253–R258. doi:10.1152/ajpregu.1990.259.2.R253. PMID 2201218.
- Bennett, A.F. (1985). "Temperature and muscle". J. Exp. Biol. 115: 333–344. PMID 3875678.
- Donley, J.M.; Shadwick, R.E.; Sepulveda, C.A.; Syme, D.A. (2007). "Thermal dependence of contractile properties of the aerobic locomotor muscle in the leopard shark and the short fin mako shark". J. Exp. Biol. 210 (7): 1194–1203. doi:10.1242/jeb.02730. PMID 17371918.
7. Nathanailides, C. (1996). Are changes in enzyme activities of fish muscle during cold acclimation significant?. Canadian Journal of Fisheries and Aquatic Sciences, 53(10), 2333-2336. http://www.nrcresearchpress.com/doi/abs/10.1139/f96-184#.Wy-tu7hZrwchttps://doi.org/10.1139/f96-184