Down syndrome research
Research of Down syndrome-related genes is based on studying the genes located on chromosome 21. In general, this leads to an overexpression of the genes.[1][2] Understanding the genes involved may help to target medical treatment to individuals with Down syndrome. It is estimated that chromosome 21 contains 200 to 250 genes.[3] Recent research has identified a region of the chromosome that contains the main genes responsible for the pathogenesis of Down syndrome,[4] located proximal to 21q22.3. The search for major genes involved in Down syndrome characteristics is normally in the region 21q21–21q22.3.

Genes
Some suspected genes involved in features of Down syndrome are given in the Table 1:
| Gene | OMIM Reference | Location | Purported Function |
|---|---|---|---|
| APP | 104760 | 21q21 | Amyloid precursor protein A4 precursor protein. Suspected to have a major role in cognitive difficulties. One of the first genes studied with transgenic mice with Down syndrome.[5] |
| SOD1 | 147450 | 21q22.1 | Superoxide dismutase. Possible role in Alzheimer's disease. Anti-oxidant as well as possible effects on the immuno-system. |
| DYRK | 600855 | 21q22.1 | Dual-specificity Tyrosine Phosphorylation-Regulated Kinase 1A. May have an effect on mental development through abnormal neurogenesis.[6] |
| IFNAR | 107450 | 21q22.1 | Interferon, Alpha, Beta, and Omega, Receptor. Responsible for the expression of interferon, which affects the immuno-system. |
| DSCR1 | 602917 | 21q22.1–21q22.2 | Down Syndrome Critical Region Gene 1. Possibly part of a signal transduction pathway involving both heart and brain.[7] |
| COL6A1 | 120220 | 21q22.3 | Collagen, type I, alpha 1 gene. May have an effect on heart disease. |
| ETS2 | 164740 | 21q22.3 | Avian Erythroblastosis Virus E26 Oncogene Homolog 2. Researchers have "demonstrated that overexpression of ETS2 results in apoptosis. Transgenic mice overexpressing ETS2 developed a smaller thymus and lymphocyte abnormalities, similar to features observed in Down syndrome."[8] ETS2-Transgenic mice were also shown to "develop neurocranial, viscerocranial and cervical skeletal abnormalities", similar skeletal abnormalities to those seen in Down's Syndrome.[9] |
| CRYA1 | 123580 | 21q22.3 | Crystallin, Alpha-A. Involved in the synthesis of Crystallin, a major component of the lens in eyes. May be cause of cataracts. |
General research
Research by Arron et al. shows that some of the phenotypes associated with Down syndrome can be related to the disregulation of transcription factors (596), and in particular, NFAT. NFAT is controlled in part by two proteins, DSCR1 and DYRK1A; these genes are located on chromosome-21 (Epstein 582). In people with Down syndrome, these proteins have 1.5 times greater concentration than normal (Arron et al. 597). The elevated levels of DSCR1 and DYRK1A keep NFAT primarily located in the cytoplasm rather than in the nucleus, preventing NFATc from activating the transcription of target genes and thus the production of certain proteins (Epstein 583).
This disregulation was discovered by testing in transgenic mice that had segments of their chromosomes duplicated to simulate a human chromosome-21 trisomy (Arron et al. 597). A test involving grip strength showed that the genetically modified mice had a significantly weaker grip, much like the characteristically poor muscle tone of an individual with Down syndrome (Arron et al. 596). The mice squeezed a probe with a paw and displayed a 0.2 newton weaker grip (Arron et al. 596). Down syndrome is also characterized by increased socialization. When modified and unmodified mice were observed for social interaction, the modified mice showed as much as 25% more interactions as compared to the unmodified mice (Arron et al. 596).
The genes that may be responsible for the phenotypes associated may be located proximal to 21q22.3. Testing by Olson and others in transgenic mice show the duplicated genes presumed to cause the phenotypes are not enough to cause the exact features. While the mice had sections of multiple genes duplicated to approximate a human chromosome-21 triplication, they only showed slight craniofacial abnormalities (688–90). The transgenic mice were compared to mice that had no gene duplication by measuring distances on various points on their skeletal structure and comparing them to the normal mice (Olson et al. 687). The exact characteristics of Down syndrome were not observed, so more genes involved for Down syndrome phenotypes have to be located elsewhere.
Reeves et al., using 250 clones of chromosome-21 and specific gene markers, were able to map the gene in mutated bacteria. The testing had 99.7% coverage of the gene with 99.9995% accuracy due to multiple redundancies in the mapping techniques. In the study 225 genes were identified (311–13).
The search for major genes that may be involved in Down syndrome symptoms is normally in the region 21q21–21q22.3. However, studies by Reeves et al. show that 41% of the genes on chromosome-21 have no functional purpose, and only 54% of functional genes have a known protein sequence. Functionality of genes was determined by a computer using exon prediction analysis (312). Exon sequence was obtained by the same procedures of the chromosome-21 mapping.
Research has led to an understanding that two genes located on chromosome-21, that code for proteins that control gene regulators, DSCR1 and DYRK1A can be responsible for some of the phenotypes associated with Down syndrome. DSCR1 and DYRK1A cannot be blamed outright for the symptoms; there are a lot of genes that have no known purpose. Much more research would be needed to produce any appropriate or ethically acceptable treatment options.
Recent use of transgenic mice to study specific genes in the Down syndrome critical region has yielded some results. APP[10] is an Amyloid beta A4 precursor protein. It is suspected to have a major role in cognitive difficulties.[11] Another gene, ETS2[12] is Avian Erythroblastosis Virus E26 Oncogene Homolog 2. Researchers have "demonstrated that over-expression of ETS2 results in apoptosis. Transgenic mice over-expressing ETS2 developed a smaller thymus and lymphocyte abnormalities, similar to features observed in Down syndrome."[12]
Specific genes
Amyloid beta (APP)
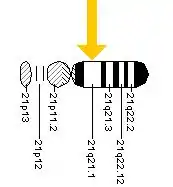
One chromosome 21 gene that might predispose Down syndrome individuals to develop Alzheimer's pathology is the gene that encodes the precursor of the amyloid protein. Neurofibrillary tangles and amyloid plaques are commonly found in both Down syndrome and Alzheimer's individuals. Layer II of the entorhinal cortex and the subiculum, both critical for memory consolidation, are among the first affected by the damage. A gradual decrease in the number of nerve cells throughout the cortex follows. A few years ago, Johns Hopkins scientists created a genetically engineered mouse called Ts65Dn (segmental trisomy 16 mouse) as an excellent model for studying the Down syndrome. Ts65Dn mouse has genes on chromosomes 16 that are very similar to the human chromosome 21 genes. Recently, researchers have used this transgenic mouse to connect APP to cognitive problems among the mice.[5]
Superoxide dismutase (SOD1)
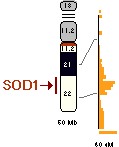
Some (but not all) studies have shown that the activity of the superoxide dismutase enzyme is elevated in Down syndrome. SOD converts oxygen radicals to hydrogen peroxide and water. Oxygen radicals produced in cells can be damaging to cellular structures, hence the important role of SOD. However, the hypothesis says that once SOD activity increases disproportionately to enzymes responsible for removal of hydrogen peroxide (e.g., glutathione peroxidase), the cells will suffer from a peroxide damage. Some scientists believe that the treatment of Down syndrome neurons with free radical scavengers can substantially prevent neuronal degeneration. Oxidative damage to neurons results in rapid brain aging similar to that of Alzheimer's disease.
Oxidative stress
The DNA oxidation product 8-OHdG is a well-established marker of oxidative DNA damage arising from oxidative stress and the excessive production of reactive oxygen species. The levels of 8-OHdG in the DNA of persons with DS measured in saliva were found to be significantly higher than in control groups.[13] 8-OHdG levels were also found to be higher in urine,[14] leukocytes[15] and fibroblasts[16] of persons with DS compared to controls. Both fetal and adult DS fibroblasts are defective in the removal of 8-OHdG as compared with age-matched cells from control healthy donors[16] These findings suggest that oxidative DNA damage may underlie some of the clinical and premature aging features of DS.
MicroRNA genes
Human chromosome 21 contains five microRNA genes: miR-99a, let-7c, miR-125b-2, miR-155, and miR-802.
Epigenetic studies of accelerated aging effects
Trisomy 21 entails an increased risk of many chronic diseases that are typically associated with older age such as an increased risk of Alzheimer's disease. The clinical manifestations of accelerated aging suggest that trisomy 21 increases the biological age of tissues, but molecular evidence for this hypothesis has been sparse. According to a biomarker of tissue age known as epigenetic clock, trisomy 21 significantly increases the age of blood and brain tissue (on average by 6.6 years).[17]
Notes
- Mao R, Zielke CL, Zielke HR, Pevsner J (May 2003). "Global up-regulation of chromosome 21 gene expression in the developing Down syndrome brain". Genomics. 81 (5): 457–67. doi:10.1016/S0888-7543(03)00035-1. PMID 12706104.
- Mao R, Wang X, Spitznagel EL, et al. (2005). "Primary and secondary transcriptional effects in the developing human Down syndrome brain and heart". Genome Biol. 6 (13): R107. doi:10.1186/gb-2005-6-13-r107. PMC 1414106. PMID 16420667.
- See Leshin, L. (2003). "Trisomy 21: The Story of Down Syndrome". Retrieved 2006-05-21.
- Rahmani Z, Blouin JL, Créau-Goldberg N, et al. (1990). "Down syndrome critical region around D21S55 on proximal 21q22.3". American Journal of Medical Genetics. Supplement. 7: 98–103. doi:10.1002/ajmg.1320370720. PMID 2149984.
- Chandra Shekhar (6 July 2006). "Down syndrome traced to one gene". The Scientist. Retrieved 2006-07-11.
- Song WJ, Sternberg LR, Kasten-Sportès C, et al. (December 1996). "Isolation of human and murine homologues of the Drosophila minibrain gene: human homologue maps to 21q22.2 in the Down syndrome "critical region"". Genomics. 38 (3): 331–9. doi:10.1006/geno.1996.0636. PMID 8975710.
- Fuentes JJ, Pritchard MA, Planas AM, Bosch A, Ferrer I, Estivill X (October 1995). "A new human gene from the Down syndrome critical region encodes a proline-rich protein highly expressed in fetal brain and heart". Hum. Mol. Genet. 4 (10): 1935–44. doi:10.1093/hmg/4.10.1935. PMID 8595418.
- Online Mendelian Inheritance in Man (OMIM): V-ETS Avian Erythroblastosis virus E26 Oncogene Homolog 2 - 164740
- Sumarsono SH, Wilson TJ, Tymms MJ, et al. (1996). "Down's Syndrome-like skeletal abnormalities in Ets2 transgenic mice". Nature. 379 (6565): 534–537. Bibcode:1996Natur.379..534H. doi:10.1038/379534a0. PMID 8596630. S2CID 4365956.
- Online Mendelian Inheritance in Man (OMIM): AMYLOID BETA A4 PRECURSOR PROTEIN; APP - 104760, gene located at 21q21. Retrieved on 2006-12-05.
- Shekhar, Chandra (2006-07-06). "Down syndrome traced to one gene". The Scientist. Retrieved 2006-07-11.
- Online Mendelian Inheritance in Man (OMIM): V-ETS AVIAN ERYTHROBLASTOSIS VIRUS E26 ONCOGENE HOMOLOG 2; ETS2 - 164740, located at 21 q22.3. Retrieved on 2006-12-05.
- Komatsu T, Duckyoung Y, Ito A, Kurosawa K, Maehata Y, Kubodera T, Ikeda M, Lee MC (September 2013). "Increased oxidative stress biomarkers in the saliva of Down syndrome patients". Arch. Oral Biol. 58 (9): 1246–50. doi:10.1016/j.archoralbio.2013.03.017. PMID 23714170.
- Jovanovic SV, Clements D, MacLeod K (December 1998). "Biomarkers of oxidative stress are significantly elevated in Down syndrome". Free Radic. Biol. Med. 25 (9): 1044–8. doi:10.1016/S0891-5849(98)00137-3. PMID 9870557.
- Pallardó FV, Degan P, d'Ischia M, Kelly FJ, Zatterale A, Calzone R, Castello G, Fernandez-Delgado R, Dunster C, Lloret A, Manini P, Pisanti MA, Vuttariello E, Pagano G (August 2006). "Multiple evidence for an early age pro-oxidant state in Down Syndrome patients". Biogerontology. 7 (4): 211–20. doi:10.1007/s10522-006-9002-5. PMID 16612664. S2CID 13657691.
- Necchi D, Pinto A, Tillhon M, Dutto I, Serafini MM, Lanni C, Govoni S, Racchi M, Prosperi E (October 2015). "Defective DNA repair and increased chromatin binding of DNA repair factors in Down syndrome fibroblasts". Mutat. Res. 780: 15–23. doi:10.1016/j.mrfmmm.2015.07.009. PMID 26258283.
- Horvath S, Garagnani P, Bacalini MG, Pirazzini C, Salvioli S, Gentilini D, Di Blasio AM, Giuliani C, Tung S, Vinters HV, Franceschi C (2015). "Accelerated epigenetic aging in Down syndrome". Aging Cell. 14 (3): 491–5. doi:10.1111/acel.12325. PMC 4406678. PMID 25678027.